Biodiversity at multiple spatial scales:
patterns and driving variables
Among-pond variability in species composition can strongly determine the total amount of aquatic pond species that occur at the landscape scale (Chase & Leibold 2002, Chase & Ryberg 2004, Williams et al. 2005). To optimize this gamma diversity, ponds should therefore be managed at the scale of pond clusters, whole landscapes or even entire biogeographic regions rather than at the scale of individual ponds. The development of efficient conservation and pond management strategies for multiple organism groups requires: (1) a good knowledge on the spatial architecture of pond biodiversity at the landscape scale; (2) a profound understanding of the mechanisms that drive gamma diversity. Both requirements imply that the contribution of each spatial scale (ranging from individual ponds, over pond clusters, groups of pond clusters and landscapes) to total national biodiversity should be identified for a wide variety of organism groups (plants and animals, poor and good dispersers, organisms with small and large body sizes).
Ponds are more isolated systems than, for example, ditches. They are also small and populations of pond organisms are therefore more prone to stochastic extinctions. As a result, the communities of many pond organisms can be strongly affected by landscape-wide meta-community processes that play far beyond the borders of individual ponds. Especially for organisms with limited dispersal abilities, isolation may impede recolonization and decrease local diversity. A lack of exchange of species between ponds may, however, also have positive effects, as it may allow communities to diverge as it impedes the spread of invasive or competitively dominant species. This may enhance among-pond diversity at the regional scale and contribute to higher gamma diversity at the scale of entire pond clusters.
Effects of land use on pond biodiversity, including ecosystem functioning, can depend on spatial scales (Chase & Leibold 2002). For example, a humped relationship between local diversity and pond primary productivity, with a decrease of alpha diversity in the upper end of the productivity spectrum was found. It is clear that such patterns are highly relevant for a better understanding of the factors that potentially determine landscape wide pond biodiversity. Chase & Ryberg (2004) also showed that the scale dependency of the relationship between pond productivity and pond biodiversity was strongly determined by the degree of isolation of the ponds, which stresses the need to consider isolation and driving environmental variables simultaneously. We will therefore combine information on connectivity (explicitly incorporated in the design of the field survey outlined in WP1) with data on local ecological characteristics, as well as with regional factors (including land use). We know from the MANSCAPE survey how these variables can efficiently and reliably be assessed (Louette et al. 2008).
Our strategy to study the spatial organization and multiscale drivers of pond biodiversity will consist of three major components:
- Analysis of extant database, in casu of the MANSCAPE database. This meta-analysis will validate results obtained for a different reason, namely to assess effect of land use at various scales on extant biodiversity. Here, we will use this database to perform a first analysis of effects of land use on scaling in diversity measures.
- Following a multi-scale survey of pond biodiversity that is specifically designed to reveal the spatial scales at which most pond biodiversity is manifested. Applying the well-established technique of ‘diversity partitioning’ (Veech et al. 2002), we will identify the spatial scales (e.g., the scale of individual ponds, the scale of pond clusters, the landscape scale) that represent the largest fraction of pond biodiversity. This exercise will allow us to develop guidelines about the spatial scale at which efforts to conserve national pond biodiversity should be focused.
- In addition to the multi-scale survey, we will carry out a detailed case study on a model system [bomb crater complex of Tommelen (Hasselt, Belgium)] that, through its extremely high degree of connectivity, may allow us to pinpoint important drivers of beta-diversity. Although the ponds all have the same age and similar size, and are located very closely to each other and share a similar environmental context (hydrology, soil, land use), they do show very pronounced differences in ecology (e.g., water turbidity, presence and development of aquatic vegetation). A detailed assessment of the environmental heterogeneity and the spatial organization of pond biodiversity in this complex may yield new insights into the relative importance of stochasticity, biotic interactions and environmental determinism as processes that shape pond communities. This is especially relevant in an effort to test how alpha, beta and gamma diversity within a pond complex can be increased by applying standard management techniques (see WP4).
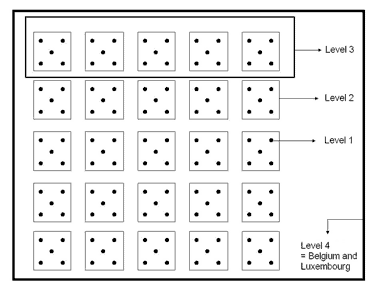
Schematic overview of the different levels of clustering applied within the multi-scale survey. This sampling design is especially created to investigate the spatial scale at which most pond biodiversity is manifested.

Aerial picture of the bomb crater field in Tommelen (Hasselt, Belgium) (© Tom De Bie).